Cell adhesion
n., plural: cell adhesions
[sɛl ədˈhiːʒən]
Definition: The binding of a cell to another cell or to an extracellular matrix component
Table of Contents
Cell Adhesion Definition
Cell adhesion is the process in which a cell uses a specialized complex of proteins to get connected and also with the substratum. Intercellular adhesion is facilitated by desmosomes, tight, and adherens junction while focal adhesion helps the cells to connect with molecules of extracellular matrix. (Portfoilo, 2022)
The adhesion of a cell to another cell or a component of the extracellular matrix is necessary for organ creation during embryogenesis and for providing a basic structure and tissue maintenance.
What is cell adhesion and why is it important?
Cell adhesion is such a mechanism by which cells adhere to themselves and with other cells with the help of cell surface proteins. The contact can either be direct like in cell junction or it can be indirect in which cells are connected by extracellular matrix adhesion. It is a gel-like material that consists of different chemicals released by the cell. Adhesion is required for cell communication and regulation, as well as for the development and maintenance of tissues. Adhesion is also required for the formation of new tissues. Cell adhesion is defined as a cell’s ability to adhere to another cell or an extracellular matrix (ECM). (Khalili & Ahmad, 2015)
What is the function of cell adhesion molecules?
Cell adhesion molecules (CAMs) are a collection of proteins on the cell surface that are necessary for the process by which cells adhere to the extracellular matrix. Adhesion cell molecules are also known as cell adhesion proteins. Cell adhesion molecules (CAMs) are molecules that assist cells to adhere to one another and their surroundings.
Cell adhesion is the binding of a cell to another cell (or a cell to an extracellular matrix component). Cell binding is essential in organ formation during embryonic development and in conferring structural framework and tissue maintenance. Without cell adhesion, the cells will be fluid as it is in certain diseases such as metastatic cancer and osteoporosis. Cell adhesion is, therefore, essential in tissue formation, angiogenesis, cell communication, and cell regulation.
Types of Cell Adhesion
Let us now know cell adhesion types. Homophilic adhesion is where cell adhesion is facilitated by the binding of similar adhesion molecules. Heterophilic adhesion – where cell adhesion is facilitated by the binding of unlike adhesion molecules.
It may also be by adhesion molecules or cells involved. Examples:
- Cadherins (cadherin cell adhesion) and nectins (nectin cell adhesion) are two well-characterized cell-cell adhesion molecules that play a role in cell adhesion regulation. (Rikitake, Mandai, & Takai, 2012)
- Epithelial cell adhesion molecule is a transmembrane glycoprotein that promotes Calcium ions homotypic cell-cell adhesion in epithelia.
- Laminate cell adhesion molecule: Laminins are a critical and physiologically active component of the basal lamina, exerting influence on proliferation and differentiation, movement, and attachment.
- Gelatin cell adhesion: Gelatin contains collagen’s RGD sequences, which makes it extremely efficient in cell-matrix adhesion.
- The neural cell adhesion molecule (NCAM) is a neuronal surface glycoprotein with immunoglobulin-like properties that interact with a range of other cell adhesion proteins to regulate adhesion, direction, and segmentation throughout neuronal growth. (Weledji & Assob, 2014)
- Fibroblast adhesion is where the adhesion of cells to the tissue matrix by molecules released by fibroblasts, e.g., fibronectin and SAM (substrate-attached material). (Harper & Juliano, 1981)
General Mechanism
Four main families of cellular adhesion molecules are selectins, immunoglobulin superfamily, integrins, and cadherins.
Cadherins and immunoglobulin superfamily are homophilic CAMs, meaning they attach to the same type of CAMs on the same cell, whereas selectins and integrins bind to various types of CAMs as they are heterophilic. Each of the adhesion molecules has a distinct function and binds with various ligands. Defects in CAM expression are frequently the cause of cell adhesion problems.
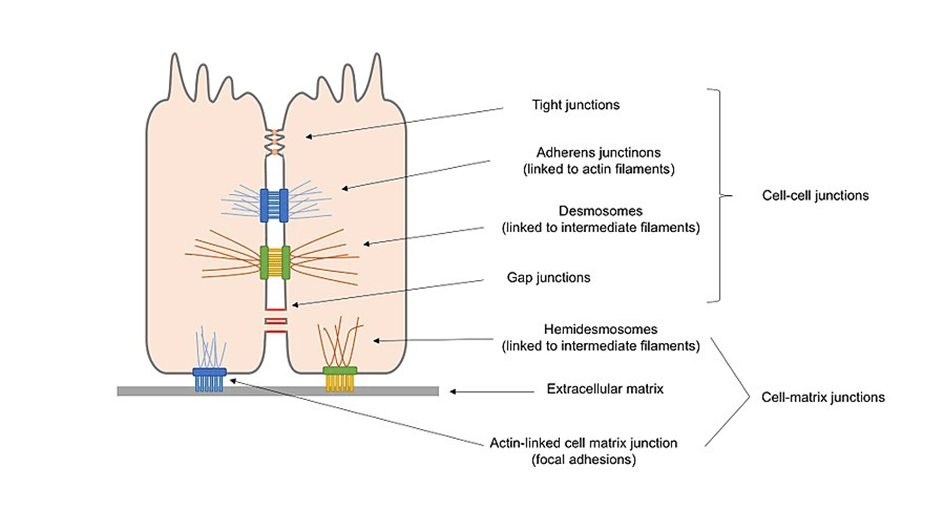
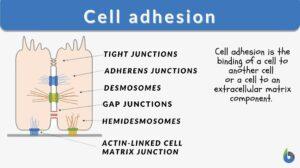
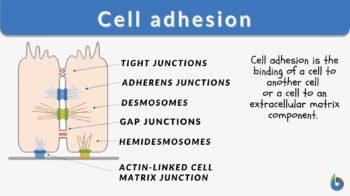
Cell junctions are formed when bindings across CAMs allow cells of multicellular organisms to adhere to one another, generating structures known as cell junctions. A second way of categorizing cell junctions is to divide them into two broad categories based on their mode of interconnecting with the cell: the first category is cell-cell junctions, which are primarily mediated by cadherins, and the second category is cell-matrix junctions, which are primarily controlled by integrins.
Following are the types of cell junctions based on their functionality.
- Anchoring junctions (desmosomes, hemidesmosomes, and adherens junctions) keep cells together as one and improve cell-to-cell interaction.
- Tight junctions (occluding junctions) are diffusion-impermeable barriers that are formed when cells come into contact.
- Gap junctions (channel-forming junctions) join nearby cells’ cytoplasms, enabling molecules to pass across.
- Signal-transmitting junctions can function similarly to synapses in the nervous system.
Cell-cell junctions
Cell-cell interactions have a variety of types. Cadherins are the most abundant CAMs identified in adherens junctions and desmosomes. This class of CAMs consists of membrane proteins with extracellular domains that promote cell-cell adhesion and operate properly when extracellular calcium ions are present. Cadherins form homophilic cell adhesion bonds between cells of the same kind, leading them to stick together. This will progress toward selective cell adhesion, which enables vertebrate cells to assemble themselves into well-organized tissues. Cadherins are proteins found in multicellular animals that are necessary for cell-cell adhesion and communication.
There are two types of cadherins:
- classical cadherins
- non-classical cadherins
Adherens junctions
Adherens junctions are largely responsible for maintaining the shape of tissues and retaining cells together. Cadherins connect cells via their cytoplasmic domain, which shares a fundamental calcium-sensitive region. When the extracellular domains of cadherins come into touch with calcium ions for homophilic interaction, they undergo a structural transition from an inactive flexible shape to a more rigid form. Cadherin intracellular domains are also highly conserved since they form complexes with catenin-like proteins.
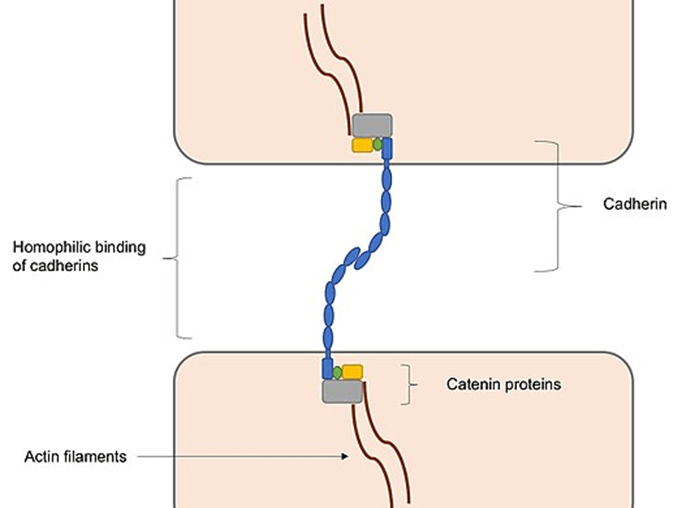
Between actin and cadherin filaments, these protein complexes behave as connective tissue. This connection with actin filaments is necessary for the development of adherens junctions, which are necessary for intracellular adhesion. Interactions with actin filaments, which are involved in the formation of adherens junctions and the creation of adhesions, may facilitate cadherin clustering. This is largely because cadherin clusters promote actin filament polymerization, which aids in the development of adherens junctions by connecting to cadherin–catenin complexes formed at the junction, hence increasing adhesion formation.
Desmosomes
Desmosomes are made up of different components for adherens junction but have an indistinguishable structure to adherens junction. Adhesion molecules include non-classical cadherins such as desmocollins and desmogleins. Rather than actin filaments, they are coupled to intermediate filaments. Because desmosomal cadherins lack catenin, their intracellular domains are associated with desmosomal plaque proteins. These proteins are involved in the formation of thick cytoplasmic adhesion plaques in desmosomes. It also attaches cadherins with the intermediate filaments. Desmosomes give rigidity and dynamic stress resistance to cells by allowing them to be released onto flexible yet elastic intermediate filaments, which is not achievable with stiff actin filaments.
Desmosomes are essential for tissues that tolerate and experience high mechanical stress. Examples of these muscles are epithelia and heart muscles, which explains their abundance in these tissues.
Tight junctions
A tight junction is a type of connection that is present in epithelial and endothelial tissues. Tight junctions serve as barriers, closing gaps and controlling the passage of molecules and extracellular fluids between cells. Tight junctions are formed when transmembrane proteins like tricellulins, occludins, and claudins bind homophilically to one another on adjacent membranes, resulting in the formation of tight junctions. To maintain the structure of the tight junction, scaffold proteins bind to the intracellular domains of these tight junction proteins, which are clustered and connected to actin filaments in a pattern similar to anchoring junctions. In addition to being essential for the creation of tight junctions, claudins also create paracellular holes that allow for the selective pathway of certain ions across tight junctions, allowing for the penetration of the membrane barrier.
Gap junctions
Connexons, which are channels, are used to construct gap junctions. Known as connexins, these are transmembrane proteins that are organized in six-membered groups on the surface of the cell membrane. Continuity channels can emerge when connexons from neighboring cells join together and align with one another. These channels not only serve to keep cells together and preserve structural stability, but they also facilitate the transport of ions and tiny molecules throughout the cytoplasms of neighboring cells.
Gap junction channels are formed as a result of the connexins that create the connexons, and they are permeable to particular ions. This allows gap junctions to participate in cell signaling by controlling the flow of molecules across signaling pathways.
Gap junctions are capable of responding in a variety of ways when subjected to a wide range of stimuli and are dynamically controlled, either through fast processes such as voltage gating or through slow mechanisms such as modifying the number of channels available in the junction.

Adhesion mediated by selectins
A family of specialized molecules known as selectins is involved in transient cell-cell adhesion in the vascular system. Selectins are found in the blood vessels. White blood cell (leukocyte) circulation in the plasma is predominantly regulated by them, with the leukocytes being able to “roll” on endothelial cells as a result of continual binding of selects. The heterophilic binding of selectins is because their extracellular domain interacts with carbohydrates on neighboring cells rather than with other selectins, and they, like cadherins, require calcium ions to function. A cell-cell adhesion between leukocytes and endothelial cells is critical for immune function because it permits leukocytes to migrate to regions of infection or injury. Activated integrins on the rolling white blood cells attach to the local endothelial cells and create a strong cell attachment, which allows the leukocytes to stop moving and go through the endothelial barrier to reach the other side.
Watch this vid on the leukocyte extravasation mechanism and the role of selectins and integrins.
Adhesion mediated by members of the immunoglobulin superfamily
One of the largest protein superfamily names is the immunoglobulin superfamily (IgSF) in the body with a large number of distinct CAMs performing a range of activities and functions. In the extracellular matrix, the transmembrane protein has one or more domains like immunoglobulin. Neural cell adhesion molecules (NCAMs) are a form of IgSF cell adhesion molecules. It forms homophilic complexes with other molecules. Heterophilic complexes are formed by vascular cell adhesion molecules (VCAMs) and intercellular cell adhesion molecules (ICAMs). Vascular endothelial cells are formed VCAMs and ICAMs. They bind with the white blood cell’s integrins and increase platelets aggregation and motility through the endothelial membrane.
Cell–matrix junctions
During the process of releasing chemicals into the extracellular space, cells create an extracellular matrix. Cells are equipped with a variety of cell adhesion molecules (CAMs), which adhere to molecules in the extracellular matrix and serve as a link between the matrix and the intracellular cytoskeleton. When linked with CAMs, the extracellular matrix can function as a framework for the organization of cells into tissues, as well as a signaling component, by activating intracellular pathways to communicate with the cells. Integrins (which cluster together like cadherins to form solid adhesions) are responsible for the regulation of the majority of cell-matrix junctions. Integrins are transmembrane heterodimers that are formed of separate subunits and have a specific domain structure, which distinguishes them from other proteins.
Both inside-out and outside-in signaling are possible for integrins. Inside-out signaling happens when the intracellular signals change the intracellular domain. They also maintain the affinity of integrin for the ligands. On the other hand, the outside-in signaling happens when the extracellular ligands get attached to the extracellular domain. It not only causes the structural change of integrins but also activates the signaling cascade. With heterophilic binding, the extracellular domain of ligands can bind with several ligands. The intracellular domain is linked with the intermediate filaments and forms hemidesmosomes.
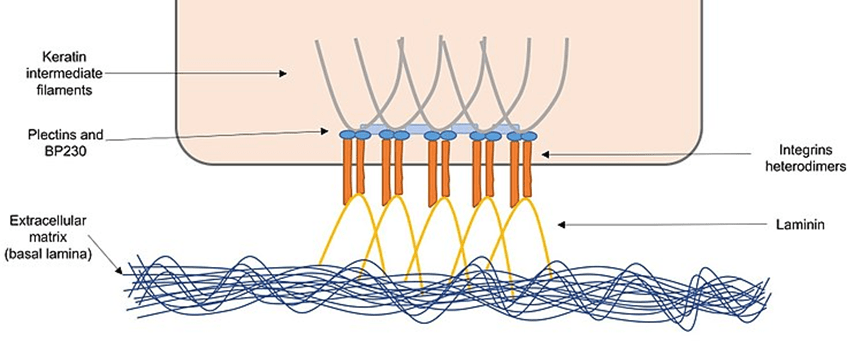
Hemidesmosomes
Integrin cell adhesion is mediated by proteins generated by epithelial cells in the extracellular matrix known as the basal lamina. Integrins use adaptor proteins like plectins and BP230 to attach the extracellular matrix to keratin intermediate filaments.
Hemidesmosomes are important for sustaining epithelial cell structural integrity by linking them indirectly through the extracellular matrix.
Focal adhesions
Integrins are proteins that link actin filaments within cells to fibronectins. Vinculins, talins, filamins and actinins are the adapter protein. These proteins form complexes at the intracellular domain, then these are formed as a fusion protein by attaching to actin filaments.
This multi-protein complex is responsible for the connection of integrins to actin filaments. For signaling complexes to operate as signals for cellular motility and development to form, they must first be formed.
Other Organisms
Eukaryotes
The evolution of cells leads to diversity among all living organisms. Plant cells are densely packed and connected with plasmodesmata. Through plasmodesmata, the molecules that may be growth or nutrients stimuli shifted either selectively or submissively towards the other plant cells.
Varieties of adhesion molecules are created by protozoans which have different and varying specificities. It connects with carbohydrates on the surface of the host cell. Adhesion between the cell helps the protozoans to adhere to and enter the host cell. The malarial parasite (Plasmodium falciparum) is an example of a hazardous protozoan. It has two adhesion molecules. The first is the circumsporozoite protein which is attached to the liver cell and the second is merozoite surface protein which gets attached to red blood cells.
To attach to host tissues or fibronectins in the extracellular matrix, the pathogenic fungus needs adhesion molecules on its cell walls. These chemicals are formed as a result of interactions between proteins or proteins and carbohydrates.
Prokaryotes
In addition to pili and flagella, prokaryotes contain adhesion molecules on their cell surface that are known as bacterial adhesins that aid in cell attachment. Adhesins are receptors that identify a variety of ligands on the cell’s surface. These molecules also have the ability to impact host specificity and regulate tropism through their interactions with different ligands (tissue- or cell-specific interactions).
Viruses
Adhesion molecules are found in viruses and are essential for viral cells to adhere to host cell membranes. For example, on the surface of the influenza virus hemagglutinin is present. It is essential for the identification of the sugar sialic acid on the host surface cell. The human immunodeficiency virus has an adhesion protein called gp120 that attaches to lymphocytes’ CD4 receptors to establish a protective barrier. To get access to host cells, the virus of hepatitis C attacks the claudins and occludins in tight junctions. This is what the virus does when it tries to get entrance to liver cells by targeting occludins and claudins in tight junctions.
Clinical Implications
Let’s take a look at the clinical implications of cell adhesion.
Adhesion in vascular injury
Thrombocyte adhesion is the initial and essential step as a result of vascular injury. In this, the platelets attach to the extracellular matrix and specific membrane receptors of the vessel and lead toward the formation of the clot. (Ruggeri & Mendolicchio, 2007)
Adhesion in inflammation
Molecules of cellular adhesion also play part in the inflammatory response. The adhesion receptors integrins, selectins, and immunoglobulin (Ig) gene superfamily regulate the several phases of white blood cells migration from the blood circulation to sites of inflammation. (González-Amaro, Diaz-González, & Sánchez-Madrid, 1998)
Additionally, there are various human genetic illnesses associated with an inability to express certain adhesion molecules. One such condition is leukocyte adhesion deficiency-I (LAD-I), which results in decreased or absent expression of the 2 integrin subunit. This results in decreased production of 2 integrin heterodimers, which are essential for white blood cells to adhere firmly to the endothelium wall in inflammatory areas to combat infections. Such patients have severe infections that can be fatal.
Platelet endothelial cell adhesion molecule, also referred to as a collection of differentiation 31 (CD31), is a protein encoded by the PECAM1 gene on chromosome 17q23.3.PECAM-1 and is involved in the removal of old neutrophils from the body. Intercellular adhesion appears to be required for neutrophil migration at sites of inflammation.
The autoimmune disease named Pemphigus is also caused by insufficient cell adhesion since it is caused by autoantibodies attacking the desmosomal cadherins of a person. It causes epidermal cells to divide and blister the skin.
To infect and cause illness, pathogenic microbes such as bacteria, viruses, and protozoans must first attach to host cells. Anti-adhesion treatment can help prevent infection. It works by attacking the adhesion molecules of the host cell or pathogen. In addition to changing adhesion molecule synthesis, competitive inhibitors that attach to adhesion molecules and hinder cell-cell interaction can be utilized as anti-adhesive medications.
Adhesion in metastasis and cancer cell
What is cell adhesion in cancer?
The inter and extracellular matrix of cancer is characterized by the presence of several diverse cell adhesion molecules. It takes several steps for cancer to develop, and adhesion molecules are essential for the production of invasive, recurrent, and distant metastasis. Evidence suggests that deviations in the adhesion properties of cancerous cells play an integral part not only in the development but also in the spread of cancer. Because intercellular adhesion loss and cell shedding from the underlying lamina propria enable the tumor tissue to depart its origin site, damage the extracellular matrix, develop a more invasive phenotype, and finally penetrate and disseminate throughout the body. (Okegawa, Pong, Li, & Hsieh, 2004)
During the process of cancer metastasis, cell adhesion failure occurs. Tumor cells that have lost their ability to adhere to one another might depart their original site of origin and multiply through the vascular system due to the breakdown of cell-cell adhesion. Cadherins are one type of CAM that is mutated in patients with cancer. They are rendered inactive either via genetic modification or through the action of other oncogenic signaling molecules. It promotes cancer cells to spread and become more metastatic as a result of the treatment. Other CAMs, such as selectins and integrins, may aid in the progression of metastasis by promoting cell-cell interaction between metastatic tumor cells that are migrating across the circulatory system. These molecules may be beneficial as cancer therapeutic targets as a result of the link between CAMs and tumor metastasis, according to the findings.
Answer the quiz below to check what you have learned so far about cell adhesion.

References
- González-Amaro, R., Diaz-González, F., & Sánchez-Madrid, F. (1998). Adhesion molecules in inflammatory diseases. Drugs, 56(6), 977-988.
- Khalili, A. A., & Ahmad, M. R. (2015). A review of cell adhesion studies for biomedical and biological applications. International journal of molecular sciences, 16(8), 18149-18184.
- Harper, P. A., & Juliano, R. L. (1981). Two distinct mechanisms of fibroblast adhesion. Nature, 290(5802), 136–138. https://doi.org/10.1038/290136a0
- Okegawa, T., Pong, R.-C., Li, Y., & Hsieh, J.-T. (2004). The role of cell adhesion molecule in cancer progression and its application in cancer therapy. Acta Biochimica Polonica, 51(2), 445-457.
- Nature Portfolio. (2022). Cell adhesion. Retrieved 18 April, 2022, from https://www.nature.com/subjects/cell-adhesion#:~:text=Cell%20adhesion%20is%20the%20process,matrix%20molecules%20through%20focal%20adhesions.
- Rikitake, Y., Mandai, K., & Takai, Y. (2012). The role of nectins in different types of cell–cell adhesion. Journal of cell science, 125(16), 3713-3722.
- Ruggeri, Z. M., & Mendolicchio, G. L. (2007). Adhesion mechanisms in platelet function. Circulation research, 100(12), 1673-1685.
- Weledji, E. P., & Assob, J. C. (2014). The ubiquitous neural cell adhesion molecule (N-CAM). Annals of medicine and surgery, 3(3), 77-81.
©BiologyOnline.com. Content provided and moderated by Biology Online Editors.