Gamete
n., plural: gametes
[ˈɡæmiːt]
Definition: A reproductive cell
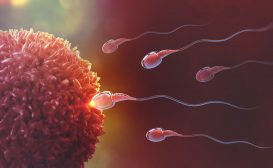
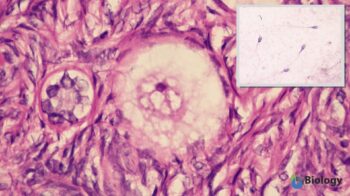
Image shows the microscopic view of human gametes, egg cell and sperm cell (inlet). Source: Modified by Maria Victoria Gonzaga, BiologyOnline.com, from the works of
Ed Uthman(human ovum – microscopy), CC BY 2.0 and Bobjgalindo (human sperm cells – microscopy), CC BY-SA 4.0 .
Table of Contents
Definition of Gamete
What is a gamete? A gamete is the mature reproductive or sex cell that contains a haploid number of chromosomes (i.e., 50% of the genetic material or only one set of dissimilar chromosomes) and is capable of fusing with another haploid reproductive cell to form a diploid zygote. The zygote is formed by the fusion (or combining) of two gametes, i.e. male gamete and female gamete. This union of gametes resulting in a zygote is called fertilization.
Characteristically, one of the gametes is usually larger in size, non-motile. It is known as female gamete or ovum or egg cell. The other gamete cell is motile and smaller in size. It is known as male gamete or sperm cell. In humans, each gamete contains 23 chromosomes and their fusion results in the formation of a diploid zygote containing 46 chromosomes. In animals, these reproductive cells are produced in respective male and female gonads or reproductive organs. In seed-bearing plants, the male gametes are the pollen while the female gametes are enclosed in the ovules of the plant. However, in plants, the gamete may or may not be always a haploid cell.
Types of Gametes
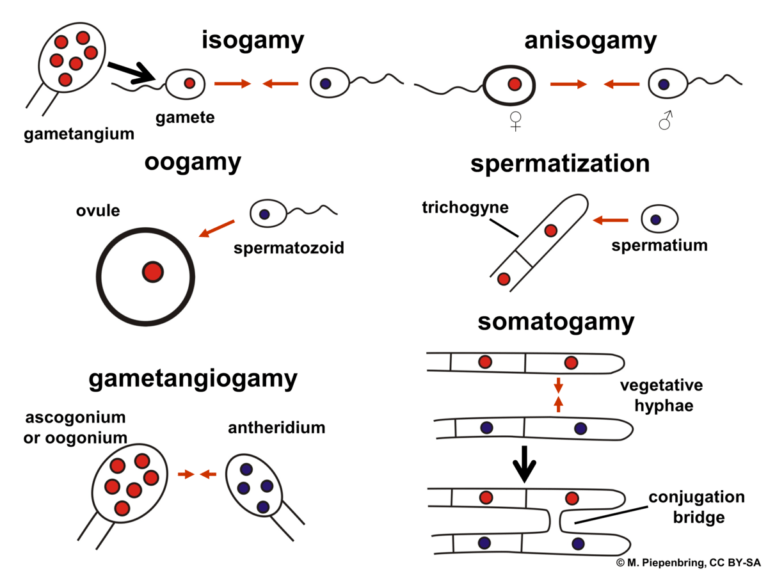
What are examples of gametes? What are the different types of gametes? The gametes involved in fertilization can be similar (referred to as isogamy) or the two gametes may be different (referred to as anisogamy).
Isogamy: gametes with similar morphology i.e., size and shape. This condition is also known as ‘heterogamy’. These types of gametes are not categorized as ‘male’ or ‘female’. These gametes are represented as ‘+’ or’-‘. Examples: gametes of unicellular algae, Chlamydomonas reinhardtii and Carteria palmata.
Anisogamy: gametes with dissimilar morphology i.e. size and shape. These types of gametes are categorized as ‘female’ and ‘male’. The gamete with a smaller size is known as sperm or male gamete whereas the gamete with a larger size is known as ova or egg or female gamete. Furthermore, these gametes can be motile as well as non-motile.
In the case of red algae, Polysiphonia, both the gametes are non-motile. The non-motile sperm fuses with the non-motile egg to form a zygote. Non-motile male gamete or sperm is known as spermatia. This is also seen in certain flowering plants wherein both the gametes are non-motile and are present in the gametophyte. The non-motile male gamete in plants is known as pollen.
Oogamy: in humans and mammals, one of the gametes, male gamete or sperm, is motile and the other gamete, egg, or female gamete is non-motile. This condition is known as oogamy, wherein a large non-motile egg is fertilized or will fuse with a small and motile sperm to form the zygote.
Another way of classification of the gametes is based on their size. Depending on the size, gametes can be categorized as:
- Microgametes: gametes that are smaller in size. These are motile, generally produced in large numbers, and don’t have storage of nutrients in them. Example: sperm cells.
- Macrogametes: gametes that are bigger or larger in size. These are non-motile, produced in limited numbers, and have storage of a large number of nutrients in them. Example: egg cells or ova.
Gamete Examples
Both males and female gametes are formed by a process known as “gametogenesis” in their respective reproductive organs. During the process of gametogenesis, a diploid (2n) cell undergoes meiosis to produce four haploid (n) cells.
In general, the process of gametogenesis usually starts with gametogonia. Gametogonia are derived from the primordial germ cells (PGCs). These germ cells proliferate by the process of mitosis. In the late embryonic stage, these cells are transferred to the gonadal ridge where they are known as gametogonia. Once gametogonia have developed, further gametogenesis will result in the formation of egg or sperm, depending on the sex of the individual.
The process of gametogenesis is completely different in males and females. The gametogenesis that results in the formation of sperm is known as “spermatogenesis” whereas the process of formation of egg or ova is known as “oogenesis”.
Structure and function – sperm cell
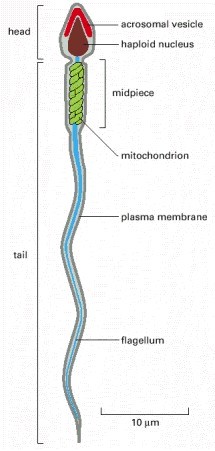
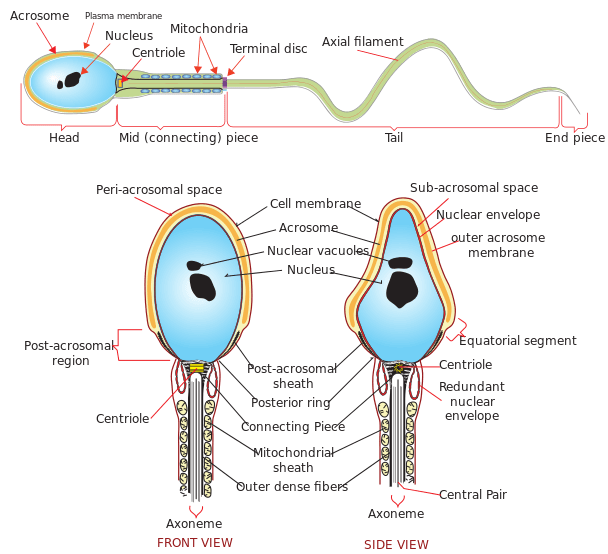
The male reproductive cell or gamete is known as the sperm cell. In animals, anisogamy is seen, wherein the structure of the male gamete is different from the female gamete. In animals, including humans, sperm is small in size and is motile. The motile organ of the sperm is known as the flagellum. The sperm cells have a limited life span and cannot divide. The sperms in mammals have two distinct structures enclosed in a single membrane (Figure 2)
- Head: contains the haploid nucleus with tightly packed DNA. Apart from the highly condensed genetic material, a sperm cell also contains a thin, flattened sac-like structure known as an acrosome and a vacuole. Acrosome contains the necessary enzyme required for breaching the ova or the egg cell. The enzyme is released by the process of exocytosis. The dimension of the head is 5.1 µm by 3.1 µm.
- Tail: drives the sperm towards the egg and eventually penetrates the egg and is located at the posterior end of the nucleus. This is also the longest part of the sperm cell. The length of the tail is 50 µm. The tail moves at a speed of 1-3 mm per minute.
Connecting the two parts, i.e. head and tail, is the neck, which is rich in mitochondria. Mitochondria is very essential for the sperm cell as it provides all the energy for the movement of the sperm. Mitochondria produces the required ATP for the movement of the sperms. Apart from the mitochondria, the neck also has centrioles. Sperm is a haploid gamete and in humans, it contains 23 chromosomes.
Spermatogenesis
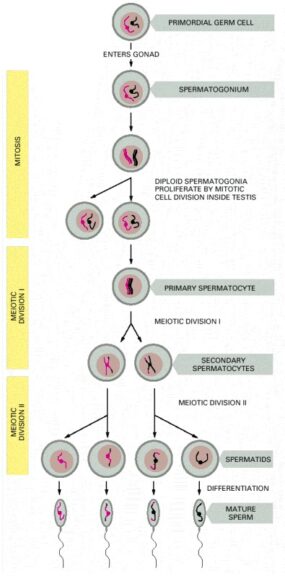
In male humans, the process of spermatogenesis occurs in the testes and is initiated only at the onset of puberty. However, once it starts, spermatogenesis is a continuous process that occurs throughout a lifetime (unlike oogenesis). Spermatogenesis occurs in the tubular structure called seminiferous tubules.
Spermatogonia, which are immature germ cells, are positioned on the outer edge of the seminiferous tubules near the basal lamina. These germ cells multiply continuously by the process of mitosis. Some of these multiplying cells do not proliferate and are converted to primary spermatocytes. These primary spermatocytes then undergo the first meiotic phase wherein, each paired homologous chromosomes contribute to cross-over and eventually undergo division I of meiosis resulting in the formation of two secondary spermatocytes, which contain 22 duplicated autosomal chromosomes (it can be a duplicated X or a duplicated Y chromosome).
These secondary spermatocytes then undergo phase II of meiosis resulting in the formation of haploid spermatids, which further undergoes differentiation to form sperms. Eventually, these sperms then move into the lumen of the seminiferous tubule. Later sperms move into a coiled tube overlying the testes known as the epididymis, where these sperms further mature and are stored in a liquid known as semen. This whole process takes around 70 days and can survive for almost 5 days in the female reproductive tract. However, outside the body, they may not survive more than a few hours.
Sperms can be stored for months or years in a frozen state and retain its capacity to fertilize eggs upon thawing. Sperm acquire energy for motility from the fructose present in the semen.
Also, an interesting fact is that sperms cannot swim backward.
Plants like, ferns, cycads and ginkgo also have flagellated sperms. Sperms of nematodes are amoeboid. They exhibit crawling movement rather than swimming.
The non-motile sperms depend on the environmental conditions for their dispersal and eventually reaching the egg cells, for example, non-motile sperms of Polysiphonia, which is a red alga, spread by water currents after released. Flies, butterflies, and insects act as a carrier for these non-motile sperms.
Sperms have acquired some major adaptations, which makes them efficient cells. Some of these adaptations are as follows:
- Streamlined structure and tapered head of sperms help them in attaining motility and agility.
- Condensed packaging of mitochondria (almost 70 in number) in the neck region of the sperms provides the energy to the sperms for their motility.
- Sperms contain some basic amines which help them to create an alkaline microenvironment even in the acidic acidic vaginal canal, thus aiding in the successful fertilization of an egg.
- The acrosome of the sperm contains lysosomal enzymes (e.g. lysozyme) that help the sperm to penetrate through the egg during the fertilization process.
The function of the sperm
The function of sperm is to reach the egg and fuse or fertilize it to form a zygote, and in that process, transfer the male genetic material and centriole (which eventually determines the microtubule cytoskeleton). The genetic material of the sperm is responsible for the traits of the progeny like the color of eyes, hair, and skin. Sperm contains X and Y chromosome that determines the sex of the progeny.
Structure and function – ovum (egg cell)
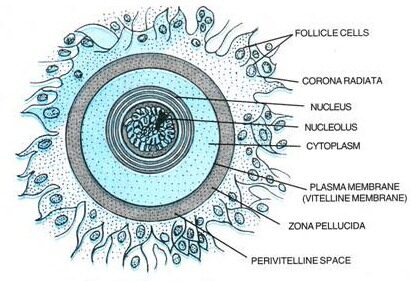
The egg cell or the ovum is the ovoid or spherical non-motile gamete produced in the female reproductive structure known as the ovaries. The size of an egg is larger than the sperm. A human egg usually has a diameter of about 0.1mm. In fish and frogs, it is around1-2mm. Ostrich egg is the largest egg — ~170 x 135 mm. Egg or ova is a haploid gamete and in humans, it contains 23 chromosomes.
Egg contains cytoplasm which is known as ooplasm. The cytoplasm of the egg has two parts i.e.., formative yolk and nutritive yolk. The human egg is described as alecithal as it contains a very small quantity of nutritive yolk. This is a contrasting feature to the eggs of birds wherein the cytoplasm is rich in nutritive yolk (which is made up of lipoproteins, pigment granules, and water). The cytoplasm contains the nucleus of the egg, known as the germinal vesicle, and a vacuole known as the germinal spot. The nucleus of the ovum is large, swollen with nucleoplasm, and is eccentric in position. Thus, giving polarity to the human ovum i.e., animal pole and vegetal pole. The animal pole is the side where the nucleus and polar body of the ovum is present, while the opposite side to it is known as the vegetal pole. The cytoplasm is enclosed in a peripheral layer known as the cortex, which is made up of microvilli and cortical granules.
Protective membranes of the ovum
The ovum is encased in a thick, transparent envelope known as the zona striata or zona pellucida. Below the zona pellucida, is a thin layer known as the vitelline membrane. There is a narrow space between the zona pellucida and the vitelline membrane which is known as perivitelline space. While the outermost layer, which is above the zona pellucida is known as the corona radiata, which radiates from the egg surface.
The function of the ova
The primary function of the ovum is to carry the genetic material, i.e. 23 sets of chromosomes in a human ovum and after fusion with the male gamete results in the formation of a zygote. It also provides the required environment that enables its fertilization by the sperm. Post-fertilization, the nutrients of the ovum are required for the growth of the zygote.
Oogenesis
Oogenesis is the process of differentiation of the ovum that occurs in the female reproductive organ known as the ovaries. The process of oogenesis differs from species to species. In humans, ova are developed from the germ cells that are present from the time of the birth of a female child. Hence, there are two broad stages of the oogenesis that occur:
- Pre-natal phase: Ova develops from the germ cell known as oogonia. The oogonia proliferate by the process of mitosis to form primary oocytes. All the germ cells proliferate to form a large number of primary oocytes at the time of birth (roughly 2 million). Post-birth, primary oocytes are not formed (this is in contrast with spermatogenesis, wherein the primary spermatocytes are continuously formed on puberty). The primary oocytes are surrounded by follicular cells and together they form a primordial follicle. The primary oocyte then initiates the first meiotic division, prior to the birth of the female child. However, this meiotic division is not completed and it halts at the prophase in the diplotene stage, until puberty. In summary, the first phase of the meiotic division is initiated in the embryo and then remains in a pause phase for around 12 years until puberty sets in. By then, it sends the signal to recommence meiosis. Interestingly, some oocytes continue to stay in the meiotic prophase for almost 50 years. Also, from the population of millions of primary oocytes, only 400 of them mature in a female’s lifetime.
- Post-natal phase: The primary oocyte remains in the latent phase until puberty. Post-birth and prior to puberty, the primary oocyte undergoes the maturation process in the ovary. During maturation, the primary oocyte remains inside the follicles wherein, they increase in size and form a membrane known as zona pellucida. Prior to ovulation, the primary oocyte resumes the meiosis and completes it. However, the cytoplasm undergoes unequal division wherein the majority of the cytoplasm is acquired by the secondary oocyte. On the other hand, the first polar body only gets a small amount of the cytoplasm. The first polar body is a small and non-functional cell that eventually degenerates. At the time of ovulation, the second meiotic division is initiated by the nucleus of the secondary oocyte; however, it progresses only up to metaphase followed by the pause in the process of the meiotic division. In case fertilization occurs, then the second meiotic division is resumed and completed.
During each menstrual cycle, the maturation of an oocyte occurs resulting in the formation of an ovum through division. This division results in cells of unequal size i.e., secondary oocytes (120-150 mm and fertilizable) and polar bodies (not more than 10 mm and non-fertilizable).
Table 1: Difference between a sperm cell and an oocyte
| Sperm cell | Oocyte |
|---|---|
| Highly motile and small in size | Non-motile and large in size |
| A very small amount of nutrients or cytoplasm and is specialized for motility | Oocyte contains a large amount of cytoplasm that provides nutrition during early development |
| There are 2 kinds of normal sperm, with respect to sex chromosomes i.e., 23, X and 23, Y | Only 1 kind of normal oocyte: 23, X |
Importance of Haploidy
Genetically, all mammals are diploids as a result of the fusion of two haploid gametes. The haploid gamete cells ensure that in every generation the genetic content or the number of the chromosomes remains constant. If the gamete cells are not haploid, then every subsequent generation will have double the chromosome number or the genetic material of the previous generation. It is important to note that a non-diploid state is the characteristic of cancer cells. The excess or a lack of a pair of chromosomes could result in destabilization during the cell replication process. This, in turn, can eventually result in a disease state, such as cancer. Ploidy alteration in humans and most mammals is usually fatal.
Apart from this, haploids are used for crop improvement (especially in rice and tobacco) as haploids can be produced in a very short span of time. Haploids are thus helpful in reducing the breeding cycle and creating novel genetic compositions for crop improvement. Haploids are also a very useful cytological tool especially for studying mutations and genetic disorders.
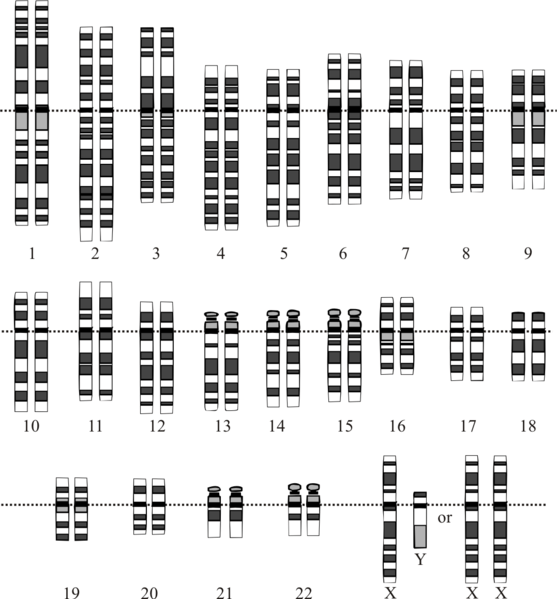
Sex Determination in Humans and other Mammals
In mammals, primary sex determination is the determination of the gonads and is determined by chromosomes and not the environment. It is the presence of a Y chromosome that determines sex in placental mammals. Normally, female cells contain two X chromosomes, i.e., XX, whereas the male cells contain an X and a Y chromosome i.e., XY. Thereby, each of the eggs from females contains single X chromosomes while the male sperms are of two types, one that contains the X chromosome and the other which contains the Y chromosome. Thus, the fusion of the egg (containing the X chromosome) with the sperm containing the X chromosome results in the formation of female progeny with XX chromosomal composition. Conversely, the male gamete containing the Y chromosome that fuses with the ovum containing the X chromosome will result in the formation of a zygote with sex chromosomes, XY, which will develop into a male offspring. The Y chromosome carries an SRY gene encoding the testis-determining factor resulting in the formation of testes in the male offspring.
Sex Determination in Birds and other Vertebrates
In birds, Z and W chromosomes are the sex determinants and the females are the ones that are heterogametic (i.e. with ZW chromosomes) as opposed to the males that are homogametic (i.e., with ZZ chromosomes). The size of the Z chromosome is larger than W chromosomes. Two genes, i.e., FET1 and ASW, present on W chromosomes determine the development of females in birds. This process of sex determination is not clear in chickens. However, like mammals, in chickens, the differentiation of gonads into male or female reproductive organs occurs after some time post-birth. Chicken needs estrogen for sex determination. Interestingly, if eggs are injected with estrogen during the developmental phase, then a male chicken can be converted to a female chicken. Some reptiles, fishes, and amphibians also exhibit ZW chromosomal sex determination.
Determination in Insects and other Invertebrates
Different insects have different patterns of sex determination. In butterflies and moths (order Lepidoptera), females are heterogametic and males are homogametic. W and Z chromosomes determine the sex in Lepidoptera. Female characteristics are associated with the W chromosome. The absence of the W chromosome results in the development of ZZ chromosomal males whereas ZO chromosomal content results in the development of females.
A moth, Talaeporia tubulosa, determines the sex, in absence of the W chromosome, by using the ambient temperature. Warm temperatures result in the formation of more female eggs and colder conditions result in a greater number of males. This is a good example of adaptation wherein certain conditions, e.g., warmth, tend to favor the formation of more female progeny as warm conditions will ensure the availability of resources for the subsequent reproduction.
In grasshoppers, XX/XO sex-determination system, which is a single-chromosome system, is used. The males possess only one sex chromosome i.e., XO (heterogametic) whereas females are XX and homogametic.
Sex determination in Drosophila
The fruit fly, Drosophila melanogaster, has been widely studied to understand heredity. In Drosophila, sex determination is based on the ratio of the number of X chromosomes to the number of sets of autosomes i.e., X: A ratio. The female-determining factors are encoded on the X chromosome whereas male-determining factors are encoded on the autosomes. The balance between X and A thus determines the sex in a fruit fly. Female flies are XX, XXY, and XXYY; male flies have XY and XO chromosomes.
Table 2: Different chromosomal content and factors that govern the sex determination in various animal species
| Female | Male | Examples | Sex determinant chromosome or factor |
|---|---|---|---|
| XX | XY | Most mammals, including humans; a few amphibians and insects | Y chromosome governs maleness |
| ZW | ZZ | birds, reptiles, amphibians | W chromosome governs femaleness |
| XX | XY | Drosophila and few insects | X:Y~ ratio > 0.75~ female X:Y~ ratio < 0.75~ male |
| XX | XO | Hemiptera (true bugs) | – |
| Diploid | Haploid | honeybees, ants | – |
| XX | XO | C. elegans | XX – hermaphrodite XO – male |
| XX, XY, YY | XX, XY, YY | Fishes | Sex determined environmentally or natural sex reversal |
Evolutionary Advantage of Gametes
Anisogamy is the evolutionary successor of isogamy. Individuals producing the same type of gametes are known as isogamous, e.g. fungi, algae, yeast. Isogamous gametes are represented as ‘+’ and ‘-‘. While in anisogamy the two gametes are morphologically different and are known as male and female. The conventional theory believes that the origin of anisogamy is based on the fact that the highest number of positive fusion of gametes occurs when the gametic material accessible for the population has undergone division with a high degree of anisogamy. It, therefore, assumes that a fixed quantity of the reserve material is essential for the zygote development and only disassortative fusions (i.e. between small and large gametes) happen. Thus, as per this theory, a large number of sperms are produced in the males to increase the fertilization probability.
Evidence suggests that fertility (i.e., the proportion of ovum that is released to that fertilized) is positively affected by the sperm density in the female tract. Hence, the higher the number of sperms in the semen, the higher is the chance of fertilization. This is also based on the fact that the higher number of sperms causes competition amongst the sperms for fertilization and therefore higher fertility. Moreover, the function of the male sperm is to transfer the genetic material only, and a large number of sperms that are small in size provide an evolutionary advantage. While the ovum invests a huge amount of energy in creating a viable zygote than a male gamete. In order to ensure a higher probability of survival, the egg contributes genetic material of its nucleus, mitochondrial genes, and provides essential nutrition for the initial development of the zygote. Thus, to provide all the necessary content, the ovum is large in size with a suitable amount of all the required content. This is thereby believed to have led to the development of anisogamy wherein eggs are non-motile, large in size and are limited in number whereas sperms are small, motile structures produced in large numbers.
Aneuploidy and Other Chromosomal anomalies in Gametes
The presence of an abnormal number of chromosomes is known as aneuploidy. A normal human cell contains 46 chromosomes. a person having 45 or 47 is aneuploid. This abnormal chromosomal number results in genetic disbalance causing a disorder. This form of mutation is the second most common type of mutations.
Aneuploidy is the result of non-separating chromosomes between two cells during cell division i.e., nondisjunction. Cases of aneuploidy in the germline cause miscarriage. A 46 XX is a normal female whereas46 XY is a normal male. The commonest form of aneuploidy is trisomy. Some known forms of aneuploidy are the following:
- Partial aneuploidy or trisomy: caused by loss or gain of part of a chromosome.
- Monosomy: lack of one chromosome of the normal complement. Turner’s syndrome is an example of monosomy. In turner’s syndrome, females have only one X chromosome, i.e. 45XO. These individuals have reduced fertility and reproductive organ growth. Turner syndrome patients are often referred to as mosaics.
- Disomy: the presence of two copies of a chromosome. It is a normal condition, however, when both copies of chromosomes are from the same parent it is known as uniparental disomy.
- Trisomy: the presence of three copies of a particular chromosome, instead of the normal two i.e presence of an extra chromosome. Trisomy of 18 is known as Edwards syndrome and trisomy of 13 is known as Patau syndrome. Trisomy of the Y chromosomes is also possible, for example (47, XXX), (47, XXY), and (47, XYY). Most of the trisomy is not viable and eventually do not survive, only a few of them are capable of surviving. The most common trisomy that is capable of surviving is Down’s syndrome. Extra chromosome 21, results in Down syndrome. It can occur in 1 in 750 births. By using prenatal screening methods such as serum screening and US surveillance almost 75% of cases are detected prior to birth. Patients with down syndrome exhibit symptoms of cognitive impairment. Another trisomy that is capable of surviving is the Klinefelter’s syndrome, wherein males have 47 chromosomes, i.e., two X chromosomes and one Y chromosome (47 XXY). These individuals have a normal life span; however, they also have low fertility and reduced development of the reproductive sex organs.
- Tetrasomy/pentasomy: Presence of four or five copies of a chromosome, respectively. Rarely seen in humans.
Dysfunctional Gametes
We’ve seen how the gametes are crucial in the perpetuation of the species through sexual reproduction and how they are essential components in promoting biodiversity (especially during the events of gamete formation and fertilization). Thus, if these gametes become dysfunctional, the species may then have to deal with reduced propagation and species diversity. Both of these are crucial in the survivability of the species. In the previous section, we’ve learned how chromosomal anomalies in humans could lead to diminished physiological functions and reproductive capacities in affected individuals. Now, let’s see the impact of dysfunctional gametes on other organisms.
In the Plasmodium life cycle
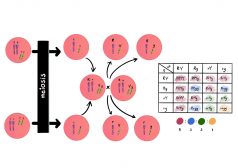
In the sexual development of the malarial parasitic protozoans, (Plasmodium spp.), the circulating female and male gametocytes undergo gametogenesis in a mosquito vector. See the figure below.
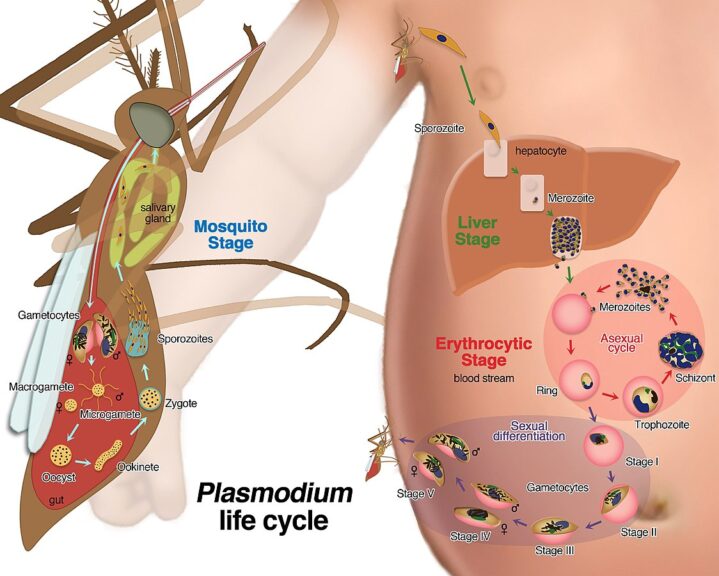
Notice how the gametocytes develop into two types: the macrogamete (female gamete) and the microgamete (male gamete). The male gamete eventually fertilizes the female gamete resulting in the formation of oocysts, which eventually develop into an ookinete.
However, the host immune factors act on these males and females throughout their sexual development and can render them dysfunctional. A dysfunctional gamete can participate in the fertilization process, however, the fusion of a healthy gamete with dysfunctional gamete results in the formation of a nonviable zygote that does not survive to reach the ookinete stage.
Try to answer the quiz below to check what you have learned so far about gametes.

References
- Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts, K., & Walter, P. (2014). Sperm. Nih.Gov; Garland Science. https://www.ncbi.nlm.nih.gov/books/NBK26914/
- Gu, N.-H., Zhao, W.-L., Wang, G.-S., & Sun, F. (2019). Comparative analysis of mammalian sperm ultrastructure reveals relationships between sperm morphology, mitochondrial functions and motility. Reproductive Biology and Endocrinology, 17(1). https://doi.org/10.1186/s12958-019-0510-y
- Lehti, M. S., & Sironen, A. (2017). Formation and function of sperm tail structures in association with sperm motility defects†. Biology of Reproduction, 97(4), 522–536. https://doi.org/10.1093/biolre/iox096
- Gilbert, S. F. (2014). Oogenesis. Nih.Gov; Sinauer Associates. https://www.ncbi.nlm.nih.gov/books/NBK10008/
- S. Georgiev (2008) Haploids in Genetic and Cytogenetical Research, Biotechnology & Biotechnological Equipment, 22:2, 644-651, DOI: 10.1080/13102818.2008.10817528.
- Gilbert, S. F. (2014). Chromosomal Sex Determination in Mammals. Nih.Gov; Sinauer Associates. https://www.ncbi.nlm.nih.gov/books/NBK9967/
- Ramiro, R. S., Alpedrinha, J., Carter, L., Gardner, A., & Reece, S. E. (2011). Sex and Death: The Effects of Innate Immune Factors on the Sexual Reproduction of Malaria Parasites. PLoS Pathogens, 7(3), e1001309. https://doi.org/10.1371/journal.ppat.1001309
- Anthony JF Griffiths, Miller, J. H., Suzuki, D. T., Lewontin, R. C., & Gelbart, W. M. (2014). Aneuploidy. Nih.Gov; W. H. Freeman. https://www.ncbi.nlm.nih.gov/books/NBK21870/
©BiologyOnline.com Content provided and moderated by Biology Online Editors.