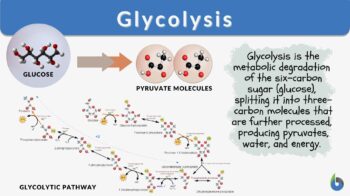
Glycolysis
n., plural: glycolyses
[ɡlaɪˈkɒlɪsɪs]
Definition: Degradation of simple sugar to pyruvic acid
Table of Contents
What is Glycolysis and Why is it Important?
Glycolysis is a metabolic pathway by which the 6-carbon molecule of glucose is broken down into a 3-C molecule called pyruvate in a series of complex oxidizing biochemical reactions. The byproduct of these reactions is the release of free energy, which is stored in the high-energy phosphate bonds of ATP and other reducing equivalents, such as NADH/H+ that are further used to produce more ATP in the mitochondria.
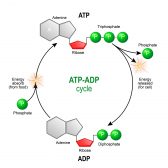
ATP is famously known as the energy currency of the cell. ATP is a high-energy molecule that fuels most of the cellular energy requirements. This is how the carbohydrates we consume in our diet are utilized inside our bodies to generate energy.
Let’s answer this common question: What is the purpose of glycolysis?
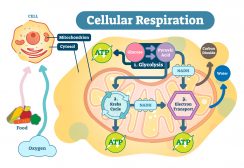
Glycolysis is an extremely important pathway that converts glucose to pyruvate. This conversion is necessary for the production of ATP without which the cells cannot survive. Glycolysis occurs in the cytoplasm of all cells. It can occur in either the presence or absence of oxygen. When it occurs in the presence of oxygen it is called aerobic glycolysis. When it occurs in the absence of oxygen it is called anaerobic glycolysis. Its presence in a wide variety of organisms suggests that it is a primitive pathway that has been the basis of the derivation of energy for a long time.
Now, let’s answer some basic questions about glycolysis:
Question: What does glycolysis produce?
Answer: Glycolysis products are 2 pyruvate, 2 NADH + 2H+ and 2ATP molecules
The net reaction of glycolysis is:
Glucose + 2 NAD+ + 2 ADP + 2 Pi → 2 pyruvates + 2 NADH + 2 H+ + 2 ATP
Question: How much NADH is produced by glycolysis?
Answer: 2 NADH molecules are produced by a single molecule of glucose in the glycolytic reaction.
Question: Where does glycolysis occur?
Answer: The location of glycolysis is the cytoplasm of the cell.
Question: What is the meaning of EMP?
Answer: EMP is an abbreviation of the Embden-Meyerhof-Parnas pathway. EMP pathway is just another name for the glycolysis pathway.
Question: Why is ATP required for glycolysis?
Answer: ATP is required in the initial few steps of glycolysis in the preparatory phase so that the 6-C glucose can be broken down into two 3-C compounds.
Question: Does glycolysis require oxygen?
Answer: Glycolysis does not necessarily require oxygen. It can occur in the absence of oxygen as well.
Biology definition:
Glycolysis is a series of reactions happening in the cytosol that results in the conversion of a monosaccharide, often glucose, into pyruvate, and the concomitant production of a relatively small amount of high-energy biomolecules, such as ATP. It is the initial metabolic pathway of cellular respiration. Cellular respiration is a series of metabolic processes wherein the biochemical energy is harvested from an organic substance (e.g. glucose) and stored in energy carriers (e.g. ATP) for use in energy-requiring activities of the cell. The major steps or processes of cellular respiration are (1) glycolysis, (2) Krebs cycle, and (3) oxidative phosphorylation.
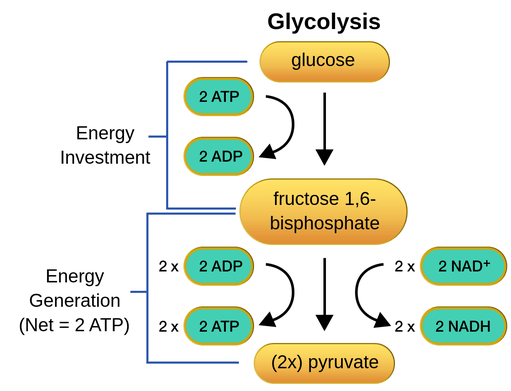
The most common and well-known type of glycolysis is the Embden-Meyerhof-Parnas pathway, which was first described by Gustav Embden, Otto Meyerhof, and Jakub Karol Parnas. Other alternative pathways are exemplified by the Entner-Doudoroff pathway and the Pentose phosphate pathway. In the Embden-Meyerhof-Parnas pathway, glycolysis is comprised of two phases: (1) the energy-investment phase (where ATP is consumed) and (2) the energy-payoff phase (where ATP is produced). The splitting of sugar during the energy-investment phase characterizes glycolysis in this regard since glucose is split into two triose phosphates: glyceraldehyde phosphate and dihydroxyacetone phosphate. The glyceraldehyde phosphate proceeds to the energy-payoff phase whereas its isomer, dihydroxyacetone phosphate, has to be converted to glyceraldehyde phosphate (via isomerase) before it can p.
Etymology: Greek “glykys”, meaning “sweet” (referring to sugar) and “Iyein”, meaning “to loosen”.
Related form: glycolytic (adj.)
See also:
History
What is the Embden–Meyerhof–Parnas (EMP) pathway? It is the most common type of glycolytic pathway which was discovered by scientists — Gustav Embden, Otto Meyerhof, and Jakub Karol Parnas. The discovery of the pathway took almost 100 years to fully unravel. The discovery of the glycolysis mechanism was originally seceded from the curiosity to understand the fermentation process, so that better wines could be made. The study of the biochemistry of glycolysis started with the experiments of the French scientist Louis Pasteur on yeast in the year 1850. He observed that the consumption of glucose decreased significantly under aerobic conditions as compared to consumption under anaerobic conditions, this effect is known as the Pasteur effect. Later, many different scientists contributed to elucidating the glycolytic pathway. Gustav Embden, by 1930 laid down all the bits and pieces of different steps together and proposed a detailed pathway.
Sequence of Reactions
Now let’s see what happens in the process of glycolysis.
Summary of reactions
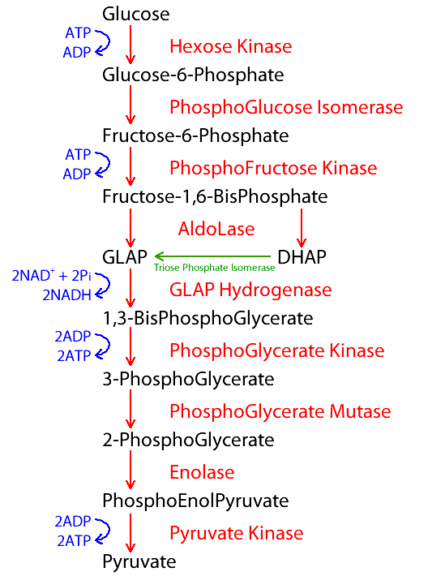
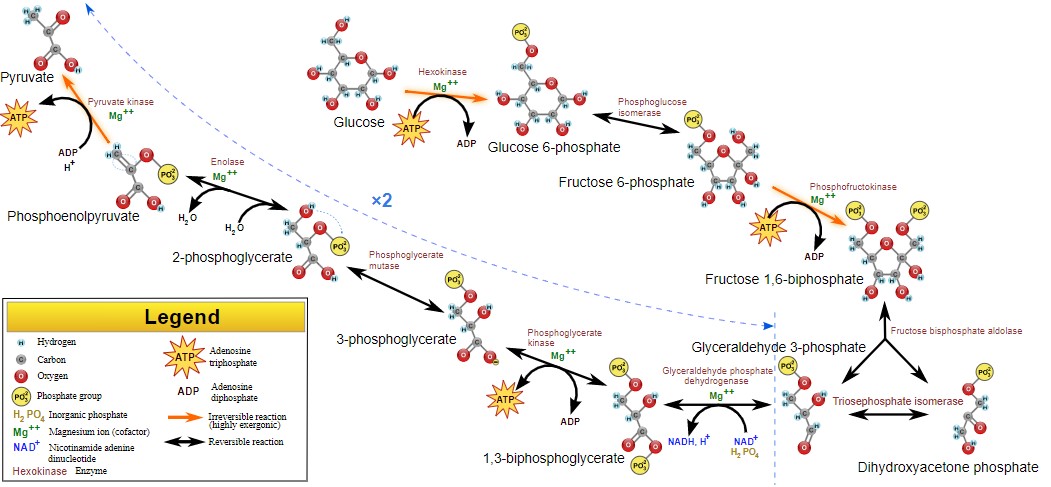
The process of glycolysis can be divided into different phases. There are a total of 9 steps of glycolysis, each of which is described below in detail.
Glycolysis equation is as follows:
Glucose + 2 NAD+ + 2 ADP + 2 Pi → 2 pyruvates + 2 NADH + 2 H+ + 2 ATP
- Preparatory phase: In this phase phosphorylation of glucose occurs followed by splitting.
- Stage of phosphorylation: Net energy is spent in this phase.
- Stage of splitting: 6-C compound splits into two 3-C compounds in this phase.
- Payoff phase: This phase is responsible for the generation of net ATP and NADH + H+

Preparatory phase: Stage of Phosphorylation
Step-1: Glucose ➔ Glucose-6-phosphate (G6P)
Glucose is phosphorylated to form Glucose-6-phosphate (G6P) by the enzyme glucokinase or hexokinase. This is an energy-requiring step, and 1 ATP is consumed for the phosphorylation of one glucose molecule. It is the first irreversible step of glycolysis. Hence, it is also known as the flux generating step. This step is important as the phosphorylated glucose molecule is trapped by the cell for further metabolism. The trapping occurs because the cell membrane is not permeable to G6P. The G6P can enter different metabolic pathways, namely Glycogen synthesis, HMP pathway, Uronic acid pathway, and Glycolysis. This article mainly focuses on the utilization of Glucose-6-phosphate in the glycolytic pathway.
Step-2: G6P ➔ Fructose-6-phosphate (F6P)
After this step, the glucose-6-phosphate is converted into fructose-6-phosphate (F6P) by the enzyme phosphohexose isomerase.
Step-3: F6P ➔ Fructose-1,6-bisphosphate (F1,6BP)
Now, the fructose-6-phosphate is once again phosphorylated by the enzyme phosphofructokinase 1 (PFK-1) to form fructose-1,6-bisphosphate (F1,6BP). This is also an energy-requiring step and consumes 1 ATP. Mg++ is a cofactor of the enzyme PFK-1. PFK-1 is also the rate-limiting enzyme of glycolysis. This step is the second irreversible step of glycolysis. It is also the first committed step of glycolysis as F1,6BP cannot escape glycolysis now. It is also known as the bottleneck of the pathway.
Preparatory phase: Stage of Splitting
Step 4: F1,6BP ➔ dihydroxyacetone phosphate (DHAP) + glyceraldehyde-3-phosphate (G3P)
F1,6BP is a 6-C compound which in this step splits into two 3-C compounds, namely, dihydroxyacetone phosphate (DHAP) and glyceraldehyde-3-phosphate (G3P) with the help of the enzyme aldolase. Phospho-triose isomerase can convert DHAP into G3P and vice versa. G3P participates in the further reactions of glycolysis.
Pay-off phase
In this phase of glycolysis, energy is yielded in the form of ATP, and reducing equivalents are generated.
Step 5: G3P ➔ 1,3 Bisphosphoglycerate (1,3BPG)
G3P molecule is converted into 1,3 Bisphosphoglycerate (1,3BPG) by the enzyme Glyceraldehyde-3-phosphate dehydrogenase. During this step, one inorganic phosphate (Pi) is utilized and one NAD+ gets reduced to NADH + H+
Step 6: 1,3BPG ➔ 3-Phosphoglycerate (3PG)
1,3 BPG is a high-energy compound, which gets converted to 3-Phosphoglycerate (3PG) by the enzyme 1,3BPG kinase. This enzyme is the only reversible kinase of the entire glycolytic pathway. In this step, substrate-level phosphorylation occurs, where ADP is converted into ATP.
Step 7: 3PG ➔ 2-phosphoglycerate (2PG)
3PG is converted into 2-phosphoglycerate (2PG) by the enzyme phosphoglycerate mutase.
Step 8: 2PG ➔ phosphoenolpyruvate (PEP)
2PG is converted into phosphoenolpyruvate (PEP) by the enzyme enolase. During this step, a water molecule is released.
Step 9: PEP ➔ pyruvate
PEP is a high-energy compound that is converted into pyruvate by the enzyme pyruvate kinase. Pyruvate is the end product of glycolysis. Second substrate-level phosphorylation occurs during this step yielding another ATP molecule.
The overall steps OR the model of glycolysis are represented by the glycolysis cycle which can be seen in this figure.
Energetics of glycolysis
The energetics of the glycolysis tells us how much ATP is produced in glycolysis. The energy produced by glycolysis is different under aerobic and anaerobic conditions. This is so because, under aerobic conditions, there is an extra generation of 2 NADH molecules that are further oxidized inside the mitochondrion to produce ATP. This oxidation of NADH within the mitochondria is responsible for the generation of 2.5 ATP molecules per NADH molecule. However, under anaerobic conditions, there is no net generation of NADH. The differences in the overall energetics of glycolysis under aerobic and anaerobic conditions are summarized in Table 1.
Table 1: Energetics of Glycolysis under aerobic and anaerobic conditions |
||
|---|---|---|
| Aerobic conditions | Anaerobic conditions | |
| Step-1 | -1 ATP | -1 ATP |
| Step-3 | -1 ATP | -1 ATP |
| Step-5 | 2 x NADH = 5 ATP | 2 x NADH = 5 ATP |
| Step-6 | 2 x ATP | 2 x ATP |
| Step-9 | 2 x ATP | 2 x ATP |
| Regeneration of NAD+ | – 2 NADH = 5 ATP | |
| NET ATP | 9-2=7 | 4-2=2 |
Biochemical logic
In the pathway of glycolysis, there are multiple regulatory steps such as Step-1 and Step-3. These different regulatory checkpoints in the pathway indicate that the intermediates of glycolysis are links to other metabolic pathways. For example, after Step-1 of glycolysis, glucose-6-phosphate can leave the glycolysis and enter the HMP pathway, glycogen synthesis, or uronic acid pathway. Similarly, after step-3, DHAP can be diverted from glycolysis by conversion into G3P. Then, G3P can enter fatty acid synthesis, or conversely when lipolysis occurs, the glycerol released can be converted into DHAP that can enter the glycolytic pathway.
Free energy changes
The free energy change is represented by ΔG. The ΔG can be calculated for each step by the equation ΔG = ΔG°’ + RTln Q, where Q represents the reaction quotient. Three steps of glycolysis are not in equilibrium as they are the irreversible steps of glycolysis. This is represented by the large free energy change of that step.
Regulation
Here are the various regulatory mechanisms involved in glycolysis.
Biological mechanisms by which enzymes are regulated
There are different sites and steps at which glycolysis is regulated by various mechanisms. The regulation of glycolysis mainly happens because of the alteration of the enzymes. The various mechanisms by which the enzymes are altered are:
- Allosteric modification – the allosteric modification is such that the substrates mostly favor the enzyme activity and products mostly inhibit the enzyme activity. So, for example, when there are more substrates, the net effect will result in the stimulation of the enzyme’s activity and favor product formation. The reverse is true when product concentration is more than the substrate.
- Gene expression
- Protein-protein interaction
- Post-translational modification
- Localization
Regulation by insulin in animals
Changing insulin and glucagon levels in the body according to the fed-fast cycle is responsible for covalent modification of the enzymes.
- When blood glucose levels are low, it stimulates the pancreatic α-cells to secrete glucagon and adrenal glands to secrete epinephrine. The Glucagon/Epinephrine in the liver phosphorylates the enzymes of glycolysis by activating protein kinase A. The enzymes of glycolysis are inactive when they are in a phosphorylated state. Hence, during a fasting state, when insulin is low and glucagon is high in the body, the enzymes of glycolysis are inactive.
- When blood glucose level is high, it stimulates pancreatic β-cells to secrete insulin. Insulin activates the enzyme phosphatase which converts the phosphorylated/active enzymes into dephosphorylated/inactive enzymes. Insulin also reduces the activity of cAMP-dependent protein kinase. Hence, under the influence of high insulin, the enzymes of glycolysis are active.
Regulation of the rate-limiting enzymes
There are four regulatory enzymes:
- Glucokinase
- Hexokinase
- Phosphofructokinase
- Pyruvate kinase
Glucokinase/Hexokinase
These two enzymes catalyze step-1 of glycolysis. The enzyme hexokinase is ubiquitous and is present in every cell whereas glucokinase is only present in certain specific cells of the liver and pancreas. Although these two enzymes catalyze the same reactions, there are certain differences between the properties of these two enzymes. The important differences between hexokinase and glucokinase enzymes are presented in Table 2 below.
Table 2: Comparison between Glucokinase and Hexokinase |
|
|---|---|
| Hexokinase | Glucokinase |
| Housekeeping enzyme – present in every cell | Inducible enzyme |
| Not induced by insulin | Induced by insulin |
| Inhibited by G6P | Not inhibited by G6P |
| High affinity for glucose – reaction is seen even in low concentrations of glucose | Low affinity for glucose – reaction is seen only with a high concentration of glucose |
| Low Km | High Km |
| No role in postprandial regulation of glucose | Role in regulation of postprandial glucose |
| Present in almost all cells | Present only in pancreas and liver |
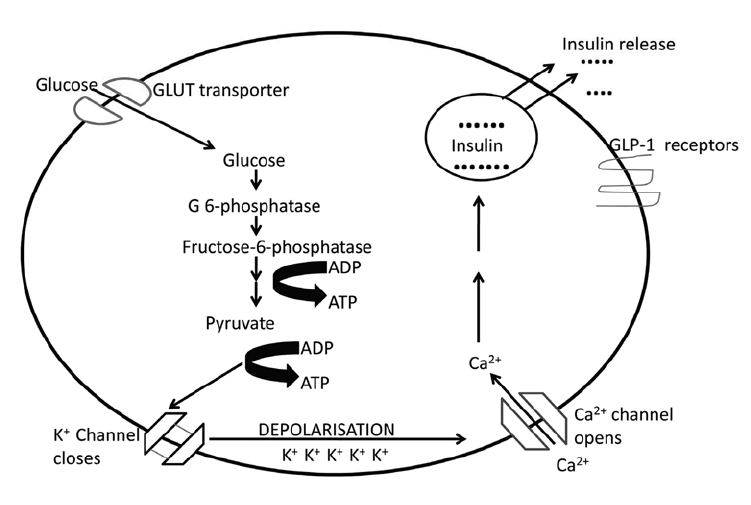
In pancreatic cells, glucokinase is active when there is abundant blood glucose. The glycolysis in the pancreatic cells produces ATP which closes the ATP-sensitive potassium channels (K+ channels). Due to the closure of K+ channels, depolarization of the cell occurs. This leads to a massive influx of calcium. Calcium is responsible for the secretion of insulin. The mechanism of insulin release can be seen in Figure 4. The insulin as previously discussed influences the activity of the glycolytic enzymes.
-
Phosphofructokinase-1 (PFK-1):
Phosphofructokinase is a very important regulatory step as it is an irreversible step of glycolysis. It has many allosteric effectors, a summary of which can be seen in Table 3. When the insulin/glucagon ratio in the blood is low, the phosphorylation/inactivation of the second phosphofructokinase occurs. This converts fructose-6-phosphate (F6P) into fructose-2,6-bisphosphate (F2,6BP). With the phosphorylation, another domain of the same enzyme F2,6BP gets activated, which converts F2,6BP back into F6P. Since F2,6BP is a very potent stimulator of the enzyme PFK-1, under low insulin/glucagon ratio, PFK-1 is inactive, and under high insulin/glucagon ratio PFK-1 is active.
When the cell energy charge is low, AMP is high. AMP is an important stimulator of PFK-1.
-
Pyruvate Kinase:
Pyruvate kinase that is in the liver is different from pyruvate kinase in muscle. In the liver, the pyruvate kinase enzyme is phosphorylated/deactivated by the epinephrine/glucagon induced protein kinase A. However, the muscle pyruvate kinase is unaffected by the epinephrine/glucagon induced protein kinase A. Hence, in a fasting state, glycolysis stops in the liver whereas it can continue to occur in the muscle.
Table 3: Glycolysis – Enzyme, Activator, and Inhibitor |
||
|---|---|---|
| Enzyme | Activator | Inhibitor |
| Hexokinase | Glucose-6-phosphate | |
| Phosphofructokinase-1 | 5’-AMP Fructose-6-phosphate Fructose-2,6-bisphosphate |
ATP Citrate Low pH |
| Pyruvate kinase | ATP | |
Post-glycolysis Processes
When glycolysis occurs, NAD+ of the cell is used up. For the glycolysis to continue there must be the regeneration of NAD+ within the cell. There are many ways by which organisms regenerate their NAD+.
Anoxic regeneration of NAD+
NAD+ regeneration, when occurs under anaerobic conditions, is called anoxic regeneration. When NAD+ regeneration occurs anaerobically, it is much faster when compared to aerobic regeneration of NAD+.
- Lactic acid fermentation – Pyruvate + NADH + H+ → lactate + NAD+
The pyruvate accepts the electrons from the NADH, this way NAD+ is regenerated, and pyruvate is converted into lactic acid. This method of regeneration of NAD+ regeneration is used by lactic acid bacteria responsible for the conversion of milk into yogurt. During intense exercise, our muscles also use lactic acid fermentation to regenerate NAD+. The pain in the muscles during intense exercise is due to the accumulation of this lactic acid.
Figure 5: Lactic acid fermentation. Image Credit: Sjantoni, CC-licensed. - Ethanol fermentation – pyruvate + NADH + H+ → ethanol + NAD+. In some yeasts, the pyruvate is first converted into acetaldehyde then later into CO2 and ethanol regenerating NAD+.
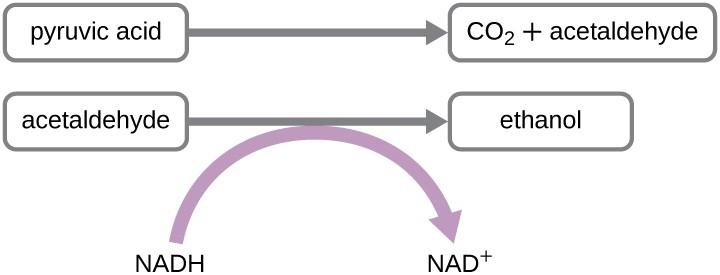
Figure 6: alcohol fermentation. Image Credit: Lumen microbiology.
Aerobic regeneration of NAD+, and disposal of pyruvate
The site of glycolysis is different from the site of oxidative phosphorylation. The ATP made during glycolysis is generated by the mitochondria. Under the aerobic condition, the NADH is transported into the mitochondria via the malate aspartate shuttle and glycerol phosphate shuttle. In the inner mitochondrial membrane, there is a sequence of electron acceptors arranged in increasing order of redox potential. The electrons are then transferred from NADH to this electron transport chain and ultimately to oxygen and thus NAD+ is regenerated. The energy released during the process is used to generate a proton gradient across the inner mitochondrial membrane. This proton gradient ultimately drives the ATP synthesis. The aerobic regeneration of NAD+ is a much more energy-efficient process than anaerobic regeneration.
The pyruvate under aerobic conditions undergoes a series of complex biochemical reactions catalyzed by a complex of three different enzymes collectively called pyruvate dehydrogenase complex (PDC). The 3-C pyruvate is converted into CO2, NADH + H+, and a 2-C acetyl CoA. The Acetyl CoA then enters the TCA cycle.
Under anaerobic conditions, the pyruvate as previously discussed gets converted into lactate or ethanol.
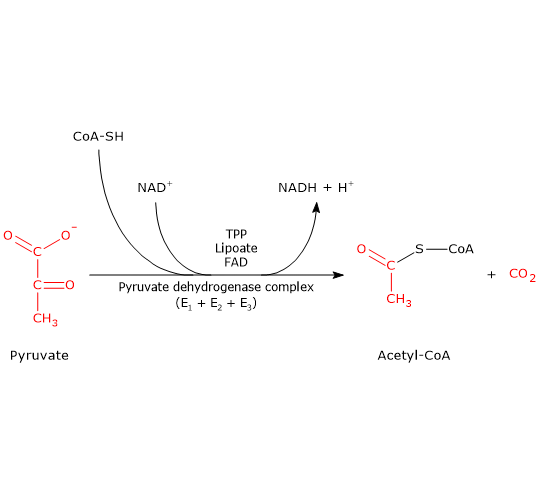
Conversion of carbohydrates into fatty acids and cholesterol
The acetyl CoA produced in the mitochondria from pyruvate is an important link between carbohydrate metabolism, fatty acid synthesis, and cholesterol synthesis. For either fatty acid synthesis or cholesterol synthesis, the acetyl CoA needs to be transported to cytosol. Since the inner mitochondrial membrane is impermeable to acetyl CoA, it uses oxaloacetate as a transporter.
Mitochondrial oxaloacetate combines with acetyl CoA to form citrate. The citrate then crosses the mitochondrial membrane to move to the cytosol. Here, citrate is cleaved by ATP-citrate lyase into acetyl CoA and oxaloacetate. The oxaloacetate inside the mitochondrion, btw, returns the acetyl CoA into the cytosol — see Figure 8. Once the acetyl CoA is in the cytosol it can either be carboxylated by acetyl CoA carboxylase enzyme to form malonyl CoA, which is the first intermediate required for fatty acid synthesis OR it can be combined with aceto-acetyl CoA to form HMG-CoA, which is the rate-limiting step of cholesterol synthesis.

Conversion of pyruvate into oxaloacetate for the citric acid cycle
After glycolysis, mitochondria handle further metabolism of pyruvate. It is oxidized by the PDC pyruvate dehydrogenase enzyme complex. The acetyl CoA thus produced enters the TCA cycle for further metabolism. The first step of the TCA cycle is the interaction between oxaloacetate and acetyl CoA to form citrate. As noted in the above figure, pyruvate can be actively converted into oxaloacetate with the help of the pyruvate carboxylase enzyme. This is an anapleurotic reaction of the TCA cycle, which means that it is a filling-up reaction that helps in increasing the other intermediates of the TCA cycle as the cycle progresses. With the increase in the intermediates of the TCA cycle, the processing capability of acetyl CoA for metabolism also increases. The key steps of the TCA cycle can be seen in Figure 9.
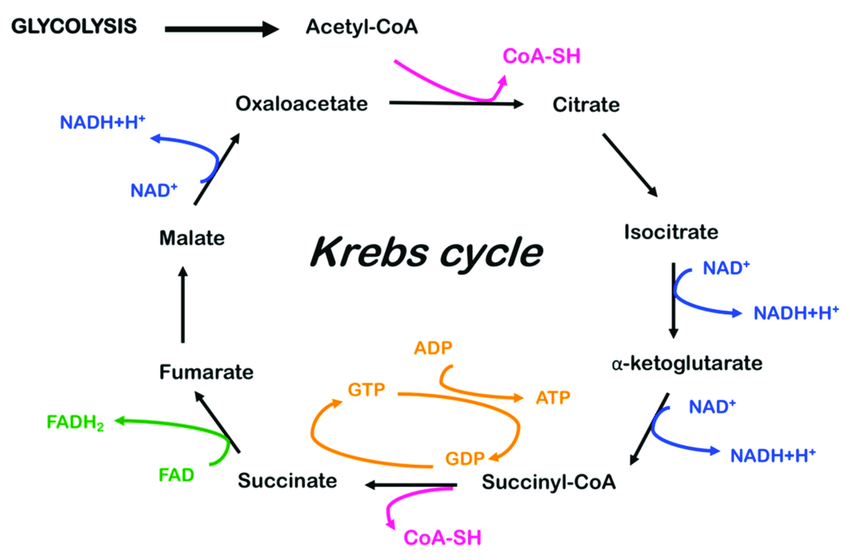
Intermediates For Other Pathways
As we already learned, a six-carbon sugar, such as glucose, is the ‘starting point’ of the glycolytic pathway. The ‘end point’ is the three-carbon molecule, pyruvate. The various products of the glycolytic pathway (referred to as glycolytic metabolites) serve as important intermediates for other metabolic pathways:
- Glucose 6-phosphate (G6P) (C6H13O9P), the first intermediate of the glycolytic pathway, may also be used as an intermediate of the Pentose Phosphate Pathway (PPP). As the name implies, this pathway produces NADPH and pentose phosphates (such as ribose 5-phosphate, which is essential in nucleotide or nucleic acid synthesis and erythrose 4-phosphate, which is for aromatic amino acid synthesis). G6P may also be stored as glycogen (such as in the liver and muscles of many animals) and as glycogen granules or intracellular starch in other organisms.
- Fructose 6-phosphate (C6H13O9P), the second intermediate of glycolysis, can also serve as an intermediate of PPP and for starch synthesis (e.g., in chloroplasts).
- Glyceraldehyde 3-phosphate (G3P) (C5H7O6P) is a glycolytic intermediate that is formed after the hexose splits into two, and through the action of the enzyme triosephosphate isomerase, G3P may be reversibly interconverted into DHAP. Apart from glycolysis, G3P is an intermediate of gluconeogenesis and also of the Calvin Cycle of photosynthesis. In photosynthesis, two molecules of G3P may be rearranged and combined to form glucose, which, in turn, may be utilized or stored as a polysaccharide (e.g., starch). G3P also has a role in the biosynthesis of thiamine (vitamin B1)
- Dihydroxyacetone phosphate (DHAP) (C5H7O6P), together with G3P, is formed after the splitting of the hexose sugar (as mentioned above). DHAP cannot enter the glycolytic pathway directly; it is first converted to G3P via the enzyme, triosephosphate isomerase. However, if not in the glycolytic pathway, DHAP enters other pathways. For instance, DHAP is used in glycogen metabolism or lipid biosynthesis. In plants, it is used in the Calvin cycle, particularly during the regeneration of RuBP (ribulose-1,5-bisphosphate).
- 1,3-biphosphoglycerate (1,3BPG) (C3H8O10P2) is the intermediate of glycolysis that is formed after the oxidation of G3P through the enzyme glyceraldehyde-3-phosphate dehydrogenase. In plants, 1,3BPG is used in the Calvin cycle, reducing it to produce G3P. In humans, about 20% of 1,3BPG produced would not go any further into the glycolytic pathway but take an alternate route, the Luebering-Rapoport pathway. In this pathway, 1,3BPG is converted into 2,3-bisphosphoglyceric acid (2,3BPG) through the action of the enzyme BPG mutase. 2,3BPG promotes the release of oxygen from the red blood cell to nearby tissues where oxygen seems more necessary.
- 3-phosphoglycerate (3PG) (C3H7O7P) is an intermediate of glycolysis formed after dephosphorylating 1,3BPG. In glycolysis, 3GP is converted into 2-phosphoglycerate by relocating the lone phosphate group. In other pathways, particularly the Calvin cycle, 3-PGA is phosphorylated to form 1,3BPG via the enzyme phosphoglycerate kinase.
- Phosphoenolpyruvate (PEP) (C3H5O6P) is an intermediate of glycolysis formed by converting 2-phosphoglycerate with the help of the enzyme enolase. In glycolysis, PEP loses the phosphate group for the final substrate-level phosphorylation and for the production of pyruvate (the final product of glycolysis) through the enzyme pyruvate kinase. In another pathway, PEP is used in the shikimate pathway to produce chorismate, which is an important intermediate in the biosynthesis of the amino acids phenylalanine, tryptophan, and tyrosine, various alkaloids, vitamin K, folate, and the plant hormone salicylic acid. In C4 plants, PEP is a substrate in the C4 carbon fixation, which is a pathway where carbon dioxide is first bound to PEP in the mesophyll cell to produce oxaloacetate. The latter is then shuttled to the bundle sheath cell where it will eventually be decarboxylated (release of carbon dioxide). The liberated carbon dioxide, in turn, is fixed via the C3 pathway. This mechanism helped C4 plants, such as sugarcane, maize, sorghum, to minimize the potential energy loss from photorespiration.
Glycolysis in Disease
Defects in the glycolytic pathway have been associated with certain diseases, such as diabetes, genetic disorders, and cancer.
Diabetes
Diabetes is a condition characterized by the inability to produce sufficient insulin. Insulin is a hormone that is crucial in maintaining the homeostatic level of glucose in the blood. Under normal conditions, the beta cells of the pancreas release an adequate amount of insulin when the blood glucose level is high. The role of insulin is to promote the uptake of glucose from the bloodstream by the cells of the body.
The inability to properly take up glucose by the cells could result in high amounts of glucose circulating in the blood. The condition wherein the blood glucose level is excessively high is referred to as hyperglycemia. Hyperglycemia is one of the symptoms of diabetes, which is a disease caused by insufficient insulin production (possibly due to a defective gene in the pancreatic beta cells involved in insulin production).
Genetic diseases
Dysfunctional genes associated with the production of enzymes involved in glycolysis could manifest as genetic disorders. Examples:
- Pyruvate kinase deficiency is a metabolic disorder caused by a mutation in the PKLR gene. This condition would therefore affect glycolysis as there would be not enough pyruvate kinase to catalyze the release of phosphate from PEP.
- Hyperinsulinemic hypoglycemia is a metabolic disorder due to a deficiency in glucokinase caused by a defective GCK gene in the pancreatic beta cells.
- Glucose-6-phosphate isomerase deficiency is a deficiency in glucose-6-phosphate isomerase (enzyme in the reaction: Glucose 6-phosphate → Fructose 6-phosphate) caused by defective GPI gene in red blood cells.
- Tarui’s disease is a deficiency in phosphofructokinase (enzyme in the reaction: β-D-fructose 6-phosphate → β-D-fructose 1,6-bisphosphate) caused by defective PFKL gene in the liver cell or by the PFKM gene in the muscle cell.
- Glycogen storage disease type XII is a metabolic disorder due to a deficiency in fructose-bisphosphate aldolase production caused by a defective ALDOA gene in the muscle, liver, and red blood cells; aldolase A is an enzyme in the reaction: Fructose 1,6-bisphosphate → Dihydroxyacetone phosphate.
- Triosephosphate isomerase deficiency, a deficiency in the triosephosphate isomerase, is caused by a defective TPI1 gene in the red blood cells. The enzyme catalyzes the reversible interconversion of G3P and DHAP.
- GSD type X is a phosphoglycerate mutase deficiency resulting in myopathy. It is caused by a defective PGAM2 gene in the muscle. The enzyme phosphoglycerate mutase is an enzyme essential in catalyzing the reaction 1,3-Bisphosphoglycerate → 3-Phosphoglycerate.
- Enolase deficiency, an autoimmune disorder due to the lack of sufficient enolase caused by a defective ENO1 gene in the red blood cells.
- Pyruvate kinase deficiency, a deficiency in pyruvate kinase, is caused by a defective PKLR gene in the red blood cells and liver.
- Baker-Winegrad disease, a deficiency in fructose bisphosphatase, is caused by a defective FBP1 gene in the liver.
Cancer
Cancer cells are essentially cells; thus, the thought that sugar consumption could make cancer “grow” faster might have some basis. By way of glycolysis, glucose, for instance, is degraded so as to produce energy via substrate-level phosphorylation. According to studies, cancer cells tend to use sugar about 200 times more than normal cells. (Ratini, 2019) Because they are a growing mass of cells, their energy requirement would apparently be high. And thus, to fuel this demand, cancer cells may utilize the glycolytic pathway more profoundly than the normal cells. Nevertheless, this topic is still undergoing a thorough study and therefore needs scientific proof to reach a wider consensus.
Try to answer the quiz below to check what you have learned so far about glycolysis.

References
- A. Hassan, Baydaa. (2019). glycolysis Embden–Meyerhof–Parnas (EMP) pathway. 10.13140/RG.2.2.36127.82089.
- Protasoni, Margherita & Zeviani, Massimo. (2021). Mitochondrial Structure and Bioenergetics in Normal and Disease Conditions. International Journal of Molecular Sciences. 22. 586. 10.3390/ijms22020586.
- Fernie, A. R., Carrari, F., & Sweetlove, L. J. (2004). Respiratory metabolism: glycolysis, the TCA cycle and mitochondrial electron transport. Current opinion in plant biology, 7(3), 254–261. https://doi.org/10.1016/j.pbi.2004.03.007
- Granner, D. K., & Rodwell, V. W. (2006). Harper’s illustrated biochemistry (27th ed.). New York: Lange Medical Books/McGraw-Hill.
- Rath, L. (2019, February 20). Cancer and Sugar: Is There a Link? WebMD; WebMD. https://www.webmd.com/cancer/features/cancer-sugar-link
©BiologyOnline.com. Content provided and moderated by Biology Online Editors.

{kind=link}
{kind=link}